


W tym rozdziale opiszemy dokładnie co leży u podstaw zjawiska uzależnienia.
Jakie procesy prowadzą do tego, że odczuwamy nieodpartą pokusę powtarzania pewnych czynności lub przyjmowania określonych substancji oraz dokonamy podziału zachowań kompulsywnych.
Słowo uzależnienie pochodzi od łacińskiego terminu „zniewolony przez” lub „związany z”. Każdy, kto usiłował przezwyciężyć jakikolwiek nałóg – lub próbował pomóc komuś innemu, aby to zrobić – na pewno rozumie dlaczego.
Uzależnienie odddziałowuje długotrwale i silnie na mózg oraz objawia się na 3 różne sposoby: silne pragnienie obiektu uzależnienia, utrata kontroli nad jego użyciem oraz ciągłe angażowanie się w nie pomimo niekorzystnych konsekwencji.
Przez wiele lat eksperci uważali, że tylko silne środki psychoaktywne mogą powodować uzależnienie. Jednak technologie neuroobrazowania i nowsze badania pokazały, że inne zachowania prowadzące do osiągania przyjemności, tj. hazard, zakupy, internet, pornografia oraz seks moga również, mówiąc w przenośni, zniewolić mózg.
Chociaż podręczniki diagnostyczne opisują wiele nałogów, z których każdy związany jest z konkretną substancją lub aktywnością, pojawia się konsensus, że mogą one reprezentować wielokrotne wyrażenia wspólnego procesu w mózgu.
Zasada przyjemności
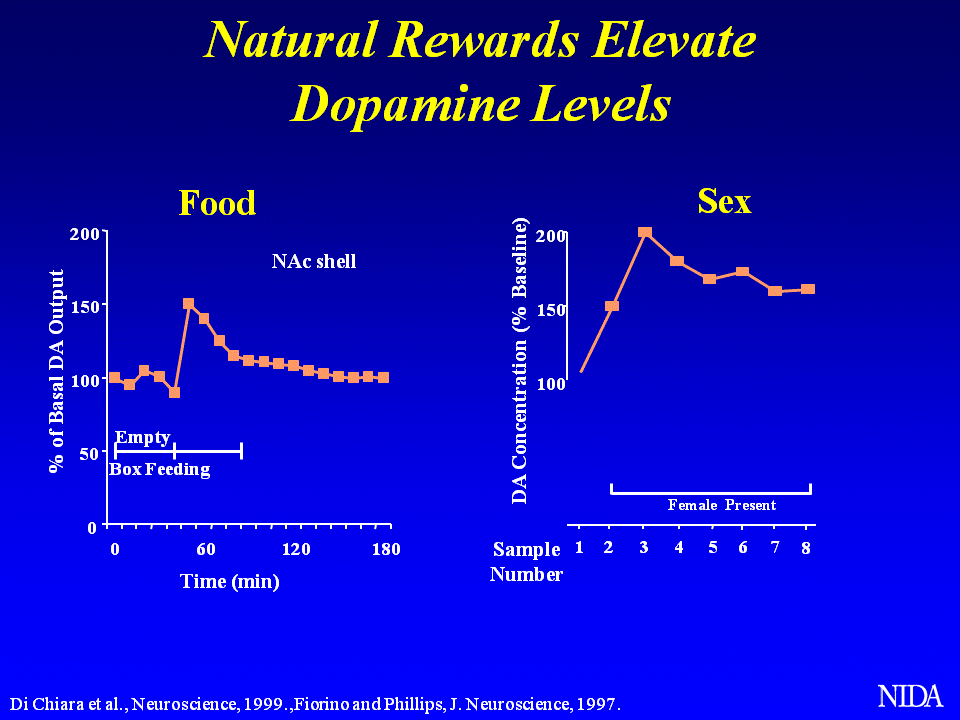
Mózg rejestruje wszystkie przyjemności w ten sam sposób, niezależnie od tego, czy pochodzą one od substancji psychoaktywnej, nagrody pieniężnej, aktu seksualnego czy satysfakcjonującego posiłku. W mózgu przyjemność ma wyraźny charakter: uwolnienie dopaminy z neuroprzekaźnika w jądrze półleżącym – skupiska komórek nerwowych leżących pod korą mózgową. Uwalnianie dopaminy powoduje aktywację receptorów dopaminowych oraz odczucie świadomej przyjemności, która pobudza hipokamp (miejsce odpowiedzialne za pamięć) oraz jądro migdałowate odpowiedzialne za emocje. Wszystko to tworzy niezwykle przyjemną mieszankę, a wspomnienia o niej wskutek pobudzenia hipokampa – jeszcze piękniejsze. Ogólna zasada jest jednak taka, że im wyższa ilość dopaminy uwalniana tym większą przyjemność odczuwamy.
Poziom dopaminy mierzony jest u szczurów umieszczając w jądrze półleżącym kaniulę, która pobiera płyn z tej części mózgu, w którym to da się określić ilość tego neuroprzekaźnika. Wzrasta ona zdecydowanie w przypadku, gdy umieścimy w klatce pożywienie lub samicę. Z doświadczenia płynie jeszcze inna obserwacja – gdy poziom głodu szczura jest niewielki, jego starania w celu uzyskania pożywienia w postaci pokonania przeszkód nie są tak intensywne jak w przypadku gdy jest wygłodniały. Wtedy zrobi niemal wszystko by zdobyć jedzenie, nawet jakby miał ryzykować utratę życia. Dzieje się tak dlatego, że działania ryzykowne są wynagradzane podwójnie przez układ nagrody, stąd też bierze się skłonność ludzi do hazardu lub uprawiania sportów ekstremalnych. Ta podwójna premia jest konsekwencją ewolucji – gdy samce idą na polowanie są przygotowane na to, że pomimo ryzyka, że mogą paść ofiarą silniejszej zwierzyny, ryzykują aby zdobyć pożywienie.
Warto zaznaczyć, że przyjmowanie tej samej substancji za pomocą różnych metod podawania może wpływać na prawdopodobieństwo doprowadzenia do uzależnienia. Palenie lub podawanie dożylne, w przeciwieństwie do przyjmowania drogą oralną, na ogół generuje szybszy i silniejszy sygnał dopaminy i jest bardziej prawdopodobne, że doprowadzi do nadużywania substancji psychoaktywnej.
Proces uczenia
Kiedyś naukowcy wierzyli, że samo doznanie przyjemności wystarcza, aby zachęcić ludzi do poszukiwania uzależniającej substancji lub jej aktywności. Ale nowsze badania sugerują, że sytuacja jest bardziej skomplikowana. Tak jak zostało wspomniane w poprzednim rozdziale, dopamina nie tylko przyczynia się do odczuwania przyjemności, ale również odgrywa wazną rolę w uczeniu się i pamięci – dwóch kluczowych elementów w przejsciu od sympatii do uzależnienia się od niej.
Zgodnie z aktualną teorią na temat uzależnień, dopamina wchodzi w interakcje z innym neuroprzekaźnikiem – glutaminianem, aby poniekąd przejąć mózgowy system uczenia się związanego z nagrodą. System ten odgrywa kluczową rolę w podtrzymywaniu życia, ponieważ łączy przyjemność i otrzymanie nagrody z działaniami niezbędnymi do przetrwania człowieka (jedzenia oraz seksu).
Układ nagrody w mózgu obejmuje obszary związane z motywacją, pamięcią oraz przyjemnością. Substancje i zachowania uzależniające stymulują ten sam obwód, a następnie go przeciążają.
Powtarzające się narażanie na uzależnieniające substancje lub zachowania powoduje, że komórki nerwowe w jądrze półleżącym i korze przedczołowej (obszary mózgu zaangażowane w planowanie i wykonywania zadań) komunikują się w taki sposób, że wywołują uczucie pragnienia, co z kolei motywuje nas do wykonania określonych działań w celu poszukiwania źródła przyjemności.
Z biegiem czasu mózg adaptuje się w sposób, który sprawia, że poszukiwana substancja lub aktywność stają się mniej przyjemne.
W naturalny sposób nagrody przychodzą zwykle z czasem i wkładem odpowiedniego wysiłku. Substancje psychoaktywne stanowią skrót, zalewając mózg dopaminą i innymi neuroprzekaźnikami. Dane pokazują, że substancje psychoaktywne mogą uwalniać od dwóch do dziesięciu razy więcej dopaminy niż naturalne nagrody, robią to szybciej i bardziej niezawodnie. U osoby uzależnionej receptory stają się przeciążone. Mózg reaguje wytwarzając mniej dopaminy lub eliminując je – adaptuje się analogicznie gdy np. zmniejszamy poziom głośności głośnika gdy jest zbyt duży hałas. W wyniku adaptacji dopamina ma mniejszy wpływ na ośrodek nagrody w mózgu. Ludzie, u których rozwija się uzależnienie często stwierdzają, że z czasem pożadana substancja nie daje im już tyle przyjemności. Muszą wziąść jej więcej, aby uzyskać ten sam efekt – ich mózgi zaadoptowały się – wystąpił efekt zwany tolerancją.
Różne substancje – ten sam efekt
Jak to możliwe, że tak różniące się od siebie substancje uzależniające, które nie mają wspólnych cech strukturalnych i wywierające różny wpływ na organizm, wywołują podobne reakcje w układzie nagrody? W jaki sposób kokaina – środek pobudzający oraz heroina – łagodzący ból środek relaksujący, będące na swój sposób tak odmienne, tam samo oddziałują na układ nagrody? Odpowiedź brzmi – wszystkie te substancje, oprócz całego wachlarza innych efektów powodują, że jądro półleżące jest zalewane dopaminą oraz nierzadko sygnałami ją mimikującymi.
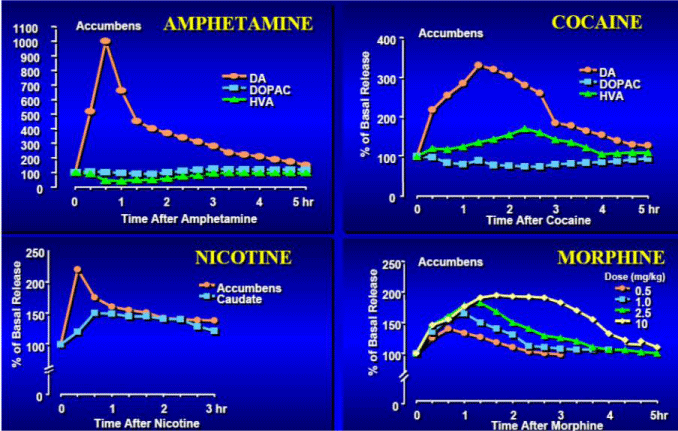
Komórka nenerwowa w VTA jest wzbudzona, wysyła wiadomość elektryczną pędzącą wzdłuż jej aksonu, będącą swojego rodzaju “autostradą” rozciągającą się do jądra półleżącego. Sygnał powoduje uwolnienie dopaminy z końcówki aksonu w maleńką przestrzeń – szczelinę synaptyczną, oddzielającą koniec aksonu od neuronu w jądrze półleżącym. Stamtąd dopamina wiąże się na swoim receptorze na neuronie półleżącym i przekazuje sygnał do komórki. Aby w kolejnym etapie wyłączyć sygnał, neuron VTA usuwa dopaminę ze szczeliny synaptycznej i “przepakowuje” ją aby ponownie użyć w razie potrzeby.
Kokaina i inne stymulanty tymczasowo wyłączają białko transportera, które zwraca neuroprzekaźnik do końcówek neuronów VTA pozostawiając w ten sposób nadmiar dopaminy działającej na jądro półleżące. Z drugiej strony heroina i inne opiaty wiążą się z neuronami w VTA, które normalnie zamykają neurony produkujące dopaminę. Opiaty uwalniają ten zacisk komórkowy, uwalniając w ten sposób komórki wydzielające dopaminę, aby wlać jej dodatkową ilość do jądra półleżącego. Opiaty mogą dodatkowo generować silny sygnał działając bezpośrednio na jądro półleżące.
Kiedy przymus przejmuje kontrolę
W pewnym momencie przyjemność związana z uzależniającą substancją lub zachowaniem ustępuje – jednak pamięć o pożądanym efekcie i potrzeba odtworzenia go (pragnienie) utrzymuje się. To tak jakby normalna machina motywacji przestała działać.
Wspomniany wcześniej proces uczenia również odgrywa tutaj znaczącą rolę. Hipokamp i ciało migdałowate przechowują informację o sygnałach środowiskowych związanych z pożądaną substancją, dzięki czemu można je ponownie zlokalizować. Te wspomnienia pomagają stworzyć warunkową reakcję – intensywne pragnienie – za każdym razem, gdy napotka się te wskazówki środowiskowe.
Żądze przyczyniają się nie tylko do uzależnienia, ale także do nawrotu po ciężko wypracowanej trzeźwości. Osoba uzaleźniona od heroiny może być zagrożona nawrotem, gdy zobaczy np.igłę do wstrzykiwań, podczas gdy inna osoba może zacząć ponownie pić po zobaczeniu butelki whisky. Uwarunkowane uczenie się pomaga wyjaśnić, dlaczego ludzie wracają do nałogów nawet po latach abstynencji.
Wczesne stadium uzależnienia charakteryzuje się tolerancją i zależnością. Sercem tej okrutnej supresji jest cząsteczka znana jako CERB (białko wiążące się z elementem odpowiedzi na cAMP). CREB jest czynnikiem transkrypcyjnym, białkiem, które reguluje ekspresję lub aktywność genów,a zatem ogólne zachowania komórek nerwowych. Gdy zażywa się addyktogeny (substancje uzależniające), wzrastają poziomy dopaminy w jądrze półleżącym, indukując komórki reagujące na dopaminę w celu zwiększenia produkcji małej cząsteczki sygnałowej – cyklicznego AMP (cAMP), która z kolei aktywuje CREB. Po włączeniu CREB wiąże się z określonym zestawem genów, uruchamiając produkcję białek kodowanych przez te geny.
Chroniczne zażywanie substancji psychoaktywnych powoduje przedłużoną aktywację CREB, która zwiększa ekspresję genów docelowych, z których część koduje białka, a następnie tłumią układ nagrody. Na przykład CERB kontroluje produkcję dynorfiny – naturalnej cząsteczki o efektach przypominających opium.
Dynorfina jest syntetyzowana przez podzbiór neuronów w jądrze półleżacym, który wraca do pętli i hamuje neurony VTA. Indukcja dynorfiny przez CERB tłumi w ten sposób obwód w układzie nagrody indukując tolerancję. Wzrost poziomów dynorfiny przyczynia się także do uzależnienia, ponieważ hamowanie ścieżki nagrody pozostawia osobę, która nie jest pod wpływem addyktogenu w depresji i niemożności czerpania przyjemności z wcześniejszych, przyjemnych czynności.
Jednak CREB to tylko mała część układanki. Ten czynnik transkrypcyjny jest wyłączany w ciągu kilku dni po zaprzestaniu zażywania substancji uzależniającej. Tak więc CREB nie może wyjaśniać długotrwałych zmian, jakie wywierają addyktogeny na mózg, powodując, że uzależnieni powracają do substancji nawet po latach czy dekadach abstynencji. Taki efekt jest w dużej mierze spowodowany sensytyzacją – zjawiskiem, które nasila działanie substancji. Dlatego też może to wydawać się sprzeczne z intuicją, ale ta sama substancja może wywołać zarówno tolerancję jak i właśnie sensytyzację.
Krótko po przyjęciu addyktogenu, aktywność CREB jest wysoka i obowiązuje reguła tolerancji– przez kilka dni użytkownik potrzebowałby większej ilości substancji aby aktywować układ nagrody. Ale jeżeli osoba uzależniona się wstrzymuje, aktywność CREB spada. W tym momencie tolerancja słabnie i pojawia się sensytyzacja, wywołująca intensywne pragnienie, które leży u podstaw kompulsywnego zachowania uzależnienia. Samo wspomnienie może przywrócić do siebie uzależnionego. Ta nieustająca tęsknota utrzymuje się nawet po długim okresie absencji. Aby zrozumieć źródła sensytyzacji, musimy jednak szukać zmian molekularnych, które trwają dłużej niż kilka dni. Jednym z winowajców jest inny czynnik transkrypcyjny: ΔFosB.
Tajemnicze ΔFosB
Coraz więcej badań wskazuje, że główny udział w uzależnieniu ma czynnik transkrypcyjny ΔFosB reprezentujący mechanizm, który wskutek zażywania środków psychoaktywnych prowadzi do zmian w mózgu, które z kolei prowadzą do uzależnienia. ΔFosB, członek rodziny czynników transkrypcyjnych Fos, gromadzi się w podzbiorze neuronów jądra półleżącego i prążkowia grzbietowego (obszary mózgu ważne dla uzależnienia) po wielokrotnym podawaniu wielu rodzajów addyktogenów. Podobna akumulacja ΔFosB występuje po kompulsywnym biegu, co sugeruje, że ΔFosB może gromadzić się w odpowiedzi na wiele rodzajów zachowań kompulsywnych. Co ważne, ΔFosB utrzymuje się w neuronach przez stosunkowo długi czas ze względu na swoją niezwykłą stabilność. Dlatego ΔFosB reprezentuje mechanizm molekularny, który może inicjować, a następnie podtrzymywać zmiany w ekspresji genów, które utrzymują się długo po ustaniu ekspozycji na daną substancję. Istnieją prace, które wskazują, że ΔFosB powoduje zwiększoną wrażliwość na behawioralne skutki nadużywania substancji, oraz być może, zwiększoną chęć ich poszukiwania. Wydaje się, że ΔFosB funkcjonuje jako rodzaj trwałego “przełącznika molekularnego“, który stopniowo przekształca ostre reakcje na daną substancję w względnie stabilne adaptacje, które przyczyniają się do długoterminowej plastyczności neuronalnej i behawioralnej, która leży u podstaw nałogu.
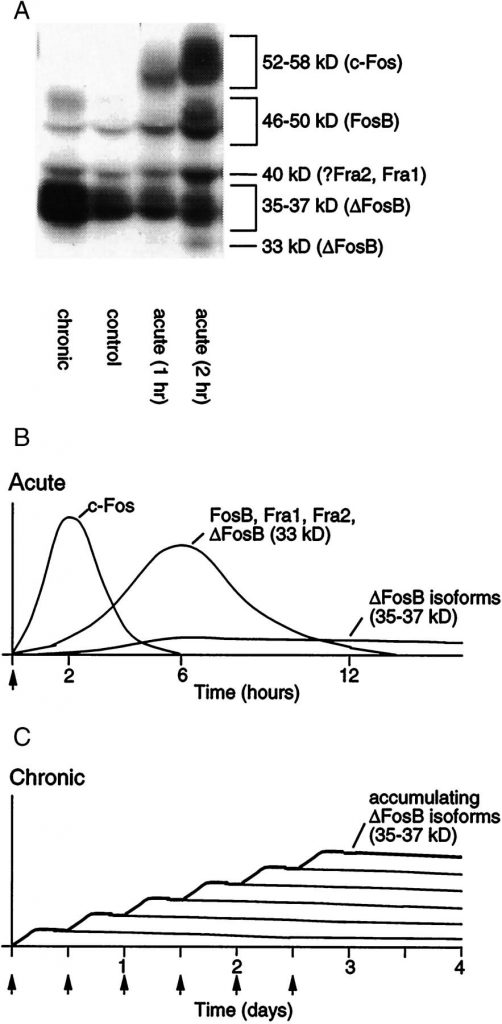
(B) Kilka fal białek podobnych do Fos [złożonych z c-Fos, FosB, ΔFosB i Fra1 lub Fra2] są indukowane w jądrze półleżącym i grzbietowych neuronach prążkowia poprzez ostre podawanie środka psychoaktywnego. Również indukowane są biochemicznie zmodyfikowane izoformy ΔFosB aczkolwiek na niskim poziomie po ostrym podaniu addyktogenu, aczkolwiek utrzymują się w mózgu przez długi czas ze względu na ich stabilność.
(C) Przy powtarzanym (np. Dwa razy dziennie) podaniu substancji, każdy ostry bodziec wywołuje niski poziom stabilnych izoform ΔFosB, co jest ukazane przez dolny zestaw zachodzących na siebie linii, które wskazują na ΔFosB indukowany przez każdy ostry bodziec. Rezultatem jest stopniowy wzrost całkowitych poziomów ΔFosB z powtarzającymi się bodźcami podczas przebiegu podawania przewlekłego, co wskazuje na zwiększającą się linie stopniową na wykresie.
W jednym z badań Eric Nestler z University of Texas wykazał, w jaki sposób uzależnienie powoduje trwałe zmiany w mózgu zwierząt. Pojedyncza dawka wielu z uzależniających substancji prowadziła do wytworzenia białka DeltaFosB, które gromadzi się w neuronach. Za każdym razem, gdy substancja jest wprowadzana do organizmu, coraz więcej DeltaFosB akumuluje się, aż zaczyna oddziaływać na genetyczny przełącznik, wpływając na to, które geny są włączone lub wyłączone. Aktywacja tego przełącznika prowadzi do zmian , które utrzymują się długo po zaprzestaniu działania substancji, prowadząc do uszkodzeń układu dopaminergicznego w mózgu, czyniąc zwierzę bardziej podatnym na uzależnienia. Ponadto zachowania inne niż przyjmowania środków psychoaktywnych, tj. bieganie czy przyjmowanie cukru, również prowadzi do akumulacji DeltaFosB i tej samej zmiany w układzie dopaminowym.
Uzależniony doświadcza pożadania, ponieważ jego mózg jest uwrażliwiony na substancję lub doświadczenie. Zjawisko to różni się o zjawiska tolerancji, gdzie uzależniony potrzebuje coraz więcej substancji , aby uzyskać ten sam przyjemny efekt. W miarę rozwoju uwrażliwienia potrzebuje on coraz mniej substancji aby jej intensywnie pożądać. Więc uwrażliwienie prowadzi do zwiększonego pragnienia, choć niekoniecznie do odczuwania przyjemnosci. To gromadzenie się DeltaFosB, spowodowane ekspozycją na uzależniającą substancję lub aktywność, prowadzi do uwrażliwienia.
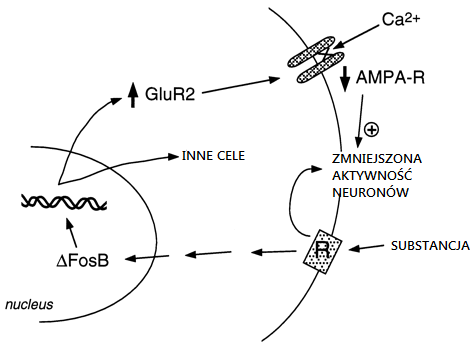
Rola ΔFosB w pośredniczeniu trwałej plastyczności na nadużywanie środków psychoaktywnych
Chociaż ΔFosB jest stosunkowo trwały, nie jest nieodwracalny. Ulega on stopniowej degradacji i nie można go już wykryć w mózgu po 1-2 miesiącach po odstawieniu środka psychoaktywnego, nawet jeśli pewne nieprawidłowości w zachowaniu utrzymują się przez znacznie dłuższy czas. Dlatego też ΔFosB per se nie wydaje się by zdolny do pośredniczenia w tych semipermanentnych nieprawidłowościach zachowania. Trudności ze znalezieniem molekularnych adaptacji, które leżą u podstaw wyjątkowo stabilnych zmian zachowań związanych z nałogiem, sa analogiczne do wyzwań stojących w obszarze uczenia się i pamięci. Chociaż istnieją niemalże idealne modele komórkowe i molekularne do tych procesów, nie udało się jak dotąd zidentyfikować molekularnych i komórkowych adaptacji, które trwają wystarczająco długo, aby mogły odpowiadać za wysoce stabilne wspomnienia behawioralne. Rzeczywiście, ΔFosB to najdłużej trwająca adaptacja jaką znamy w dorosłym mózgu, powstająca nie tylko w odpowiedzi na nadużywanie środków psychoaktywnych, ale także inne zaburzenia (nie obejmujące zmian chorobowych). W związku z tą rozbieżnością rozwinęły się dwie propozycje, zarówno w dziedzinie uzależnień, jak i uczenia się i pamięci.
Teraz proponuję uzbroić się w cierpliwość gdyż kolejne 2 akapity są dość ciężkostrawne i nie każdy może je zrozumieć.
Jeżeli masz niską tolerancję na typowo naukowy język możesz je opuścić. Jednak aby artykuł był kompletny, uważam że niezbędnym jest wyjaśnienie poniższego problemu. Później będzie już tylko z górki 😉
Jedną z możliwości jest to, że bardziej przejściowe zmiany w ekspresji genów, takie jak pośredniczone przez ΔFosB lub inne czynniki transkrypcyjne (np. CREB) mogą pośredniczyć w bardziej długotrwałych zmianach w morfologii neuronów i strukturze synaptycznej. Na przykład – zwiększenie gęstości kolców dendrycznych (szczególnie zwiększenie dwugłowych grzbietów) towarzyszy zwiększonej skuteczności synaps glutaminergicznych w neuronach piramidalnych hipokampa podczas długotrwałego wzmacniania i odpowiada zwiększonej wrażliwości behawioralnej na kokainę na poziomie średnich neuronów kolczastych jądra półleżącego. Nie wiadomo, czy takie zmiany strukturalne są wystarczająco długotrwałe, aby uwzględnić wysoce stabilne zmiany w zachowaniu, chociaż te ostatnie utrzymują się przez co najmniej 1 miesiąc od odstawienia addyktogenu. Ostatnie dowody wskazują na możliwość, że ΔFosB i jego indukcja Cdk5 jest jednym z mediatorów zmian w strukturze synaptycznej w jądrze półleżącym. Tym samym infuzja inhibitora Cdk5 do jądra półleżącego zapobiega zdolności wielokrotnej ekspozycji na kokainę w celu zwiększenia gęstości dendrytycznej kręgosłupa w tym regionie. Jest to zgodne z poglądem, że Cdk5 który obficie występuje w mózgu, reguluje strukturę nerwową i wzrost. Jest to możliwe, choć jak do tej pory nieudowodnione, że takie zmiany w morfologii neuronów mogą przetrwać sam sygnał ΔFosB.
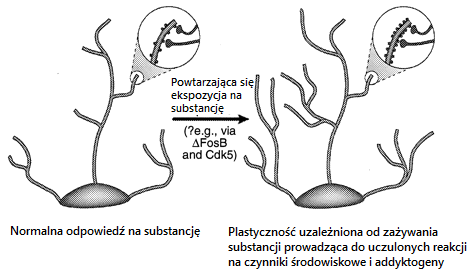
Inna możliwość jest taka,że przejściowa indukcja czynnika transkrypcyjnego (ΔFosB, CREB) prowadzi do bardziej trwałych zmian ekspresji genów poprzez modyfikację chromatyny. Uważa się, że te i wiele innych czynników transkrypcyjnych aktywuje lub tłumi transkrypcję docelowego genu poprzez promowanie odpowiednio acetylacji lub deacetylacji histonów pobliżu genu. Chociaż takie acetylowanie i deacetylacja histonów mogą występować bardzo szybko, możliwe jest, że ΔFosB lub CREB może powodować dłuższe adaptacje machiny enzymatycznej kontrolującej acetylację histonów. ΔFosB lub CREB może również promować dłuższe zmiany w ekspresji genów poprzez regulację innych modyfikacji chromatyny (np. metylacja DNA lub histonów), które są zaangażowane w trwałe zmiany w transkrypcji genów. Chociaż możliwość ta jest ciągle w sferze spekulacji, może ona dawać pewien pogląd na mechanizm, dzięki któremu przemijające adaptacje na substancję nadużywającą prowadzi do zasadniczo trwałych konsekwencji behawioralnych.
“Supernormal stimuli”
Niektóre badania sugerują, że pewne rzeczy, które dzisiaj lubimy, zostałyby sklasyfikowane jako “bodźce nadprzyrodzone“, określane przez biologów ewolucyjnych jako opisujące każdy bodziec, który wywołuje reakcję silniejszą niż bodziec, dla którego ewoluował, nawet jeśli jest sztuczny.
Nikolaas Tinbergen, laureat nagrody Nobla, jest ojcem terminu “nadprzyrodzone bodźce”. Przeprowadził szereg eksperymentów mających na celu wyjaśnienie tego zjawiska. W jednym z nich skonstruował jaja z gipsu, aby zobaczyć na których ptak będzie preferował siedzieć. Okazało się, że wybierał chętniej te, które były większe, miały specyficzne oznaczenia i były bardziej nasycone kolorem.
Odkrył również, że terytorialna samcza ryba z gatunku stickleback zaatakowała drewniany model ryby bardziej energicznie niż byłby to prawdziwy samiec. A jeszcze tym agresywniej, im bardziej spód manekina był bardziej czerwony.
Zbudował również tekturowe imitacje samic motyli z bardziej zdefiniowanymi znakami, którymi w bardzo szybkim tempie był w stanie wpływać na zachowanie tych zwierząt za pomocą nowego “super bodźca”, którego samce preferowały bardziej od naturalnych samic.
Instynkt przejął kontrolę, a ich zachowania stawały się szkodliwe dla ich przetrwania, gdyż nie mogły sobie odmówić reakcji na fałszywy bodziec.
Wiele prac Tinbergena zostało ciekawie opisanych przz psychologa z Harvardu Deirdre’a Barretta w książce “Supernormal Stimuli: How Primal Urges Overran Their Evolutionary Purpose”. Warto by się zastanowić, czy przełożenie tych odkryć do ludzkich zachowań jest bliskie czy dalekie. Dr Barrett wydaje się myśleć, że związek jest bliższy niż sądzimy, argumentując, że stymulacja sztucznymi bodźcami rządzi zachowaniami ludzi równie silnie jak zwierzętami. Hipoteza mówi, że podobnie jak szybkie wprowadzenie przez Tinbergena nadmiernej stymulacji u zwierząt wywołało niepokojące efekty, postępująca technologia może stworzyć analogiczną sytuację u ludzi.
Wysoce uzależniający charakter fast-foodów jest jednym z największych problemów naszego pokolenia. Zostało ono zaprojektowane tak, aby było bardziej atrakcyjne niż jego naturalne odpowiedniki. Czy można się zatem dziwić, że kiedy zostaje on wprowadzany do kolejnych krajów, jego konsumpcja natychmiastowo wzrasta?
Przez długi okres czasu paleta naszego pożywienia utrzymywała się na w miarę stałym poziomie. Aktualnie praktycznie co miesiąc mamy jakieś nowe mieszanki żywnościowe. W jaki sposób wpływa to na nasze zdrowie możemy tylko spekulować. Warto zapamiętać, że jakość żywności jest jedną z najtrudniejszych rzeczy, którą można kontrolować, gdyż jedzenie jest absolutną koniecznością, a fast-food stanowi “super stymulującą” wersję naturalnej nagrody, którą powinniśmy realizować. Uzależnienie od jedzenia to prawdziwe wyzwanie, gdyż potencjalne miejsca, gdzie możemy ulec pokusie są wszechobecne.
Gry video i telewizja
Uzależnienie od telewizji może powodować, że niektórzy użytkownicy przejawiają oznaki uzależnienia od zachowań. Ludzie często oglądają telewizję, by zmienić nastrój, jednak ulga jaką otrzymują jest chwilowa i często prowadzi do chęci powtórzenia tej czynności.
Nie jest z kolei tajemnicą, że gry komputerowe powiązane są z eskapizmem, ale mało kto wie, że niektóre badania wskazują na pewne objawy odstawienia – huśtawki nastrojów, nadpobudliwość a nawet objawy fizyczne.
Internet
Jak można się spodziewać, psychologowie poważnie koncentrują się tematyce sieci, gdyż może ona stanowić bardzo niebezpieczne źródło uzależnienia. Internet pozwala nieskrępowanie i poza kontrolą angażować się w niemalże wszystko. Prowadzi to do wielu problemów, np. Japonia i Korea Południowa mają spore trudności z wieloma ludźmi o podatnych osobowościach, które mają niezdrową obsesję na punkcie internetu. Jedna historia opowiada o człowieku, który nie opuścił swojego mieszkania przez pół roku przegladając internet.
Dodatkowo media społecznościowe powodują, że niektórzy ludzie sa bardziej przygnębieni oglądając innych prywatne życia po drugiej stronie ekranu, które (najczęściej tylko w internecie) mogą wyglądać na bardziej atrakcyjne od nich. Jednak mimo to nie mogą przestać sprawdzać, myśląc że może coś ich ominąć.
Ponadto nadużywanie internetu dla niektórych moze oznaczać poważne problemy z umiejętnością koncentracji uwagi. Szybka rozrywka, która zapewnia przeglądanie sieci oraz fakt, że informacje dostępne są na “jedno kliknięcie”, może prowadzić do spadku myślenia koncepcyjnego i krytycznego. Niektórzy wręcz twierdzą, że internet może stać się “chronicznym dystraktorem”, który powoli pożera cierpliwość i umiejętność myślenia oraz pracy nad rzeczami przez dłuższy czas.
Pod tym linkiem znajdziecie ponad 100 opracowań naukowych dotyczących hipofrontalności (zmienionego funkcjonowania przedczołowego) u osób uzależnionych od internetu
Hazard
Dotychczasowe badania pokazują, że hazardziści i osoby nadużywające środków psychoaktywnych mają wiele wspólnych genetycznych predyspozycji do impulsywności oraz poszukiwania nagrody. Tak jak uzależnieni od substancji wymagają coraz to większych dawek aby osiągnąć pożądany poziom przyjemności, kompulsywni hazardziści podejmują coraz bardziej ryzykowne przedsięwzięcia. Analogicznie do osób nadużywających addyktogenów – hazardziści odczuwają również objawy odstawienia, gdy są relatywnie długo odseparowani od możliwości dostarczenia “dreszczyku” emocji”.
Ponadto w trakcie badań okazało się, że podobnie jak środki psychoaktywne, hazard aktywuje podobne regiony w mózgu w analogiczny sposób. Wyniki te pochodzą z obserwacji przepływu krwi i aktywności elektrycznej w mózgach ludzi wykonujących różne zadania przy komputerach, które albo naśladują grę w kasynie, albo sprawdzają kontrolę impulsów. W niektórych eksperymentach karty wirtualne wybrane z różnych talii zarabiają lub tracą pieniądze graczy, inne zadania wymagają od badanych szybkiego reagowania na niektóre obrazy, które migają na ekranie, jednocześnie nie reagując na inne.
Wyniki badań przeprowadzonych w 2005 roku z wykorzystaniem gry karcianej sugerują, że osoby uzależnione od hazardu, podobnie jak uzależnione od addyktogenów, tracili wrażliwość w odpowiedzi na bodziec – podczas wygrywania mieli niższą aktywność elektryczną w kluczowym regionie układu nagrody. W badaniu z 2003 roku na Uniwersytecie w Yale oraz badaniu przeprowadzonym w 2012 roku na Uniwersytecie w Amsterdamie gracze nałogowi, którzy brali udział w testach mierzących swoją impulsywność, mieli niezwykle niski poziom aktywności elektrycznej w rejonach mózgu przedczołowych, które normalnie odpowiadają za ocenę ryzyka i tłumienie instynktów.
Co ciekawe – na przestrzeni dziesięcioleci naukowcy zauważyli, że niezwykle duża liczba pacjentów z chorobą Parkinsona – od 2 do 7 procent – jest kompulsywnymi hazardzistami. Leczenie jednego zaburzenia najprawdopodobniej przyczynia się do powstawania drugiego. Aby złagodzić objawy Parkinsona, pacjenci przyjmują lewodopę i inne leki zwiększające poziom dopaminy. Naukowcy sądzą, że w niektórych przypadkach napływ neuroprzekaźnika modyfikuje mózg w sposób, który stwarza większe ryzyko podejmowania ryzykownych decyzji np. w grze pokera.
Pornografia
Prawdopodobnie najbardziej kontrowersyjny ze wszystkich współczesnych bodźców, pornografia została opisana jako podstępna w naturze, ponieważ mogłaby wypaczać normalną aktywność seksualną. Pornografię powiązano ze zmieniającymi się gustami seksualnymi, a niektórzy wręcz twierdzą, że porno może stać się niekończącym się wyzwalaczem dopaminy (mimo że istnieje niewiele badań przeprowadzonych na porno i umyśle).
Istnieje fragment powieści Kurta Vonneguta, w którym mężczyzna pokazuje innemu mężczyźnie fotografię kobiety w bikini i pyta: “I jak Ci się podoba ta dziewczyna?”. Po czym odpowiedź mężczyzny brzmi: “To nie jest dziewczyna. To jest kartka papieru.” Ci, którzy ostrzegają przed uzależniającą naturą porno, zawsze podkreślają, że nie jest ona uzależnieniem seksualnym, tylko technologicznym. Sugeruje się, że pornografia zakłóca “układ nagrody” w ludzkiej seksualności. Po co próbować zaimponować potencjalnemu partnerowi, jeżeli możesz po prostu pójść do domu i spojrzeć na porno? Uznaje się to za początek uzależnienia od pornografii, poniewaz nowość jest zawsze “na jedno kliknięcie”, a jest ona ściśle związana z wysoce uzależniającą naturą dopaminy.
Zarówno doświadczenia seksualne, jak i substancje działają poprzez receptory D1 w nucleus accumbens. Nowość seksualna jest fascynująca, ponieważ wyzwala wybuchy dopaminy w regionach mózgu silnie związanych z nagradzaniem i ukierunkowanymi na cel zachowaniami. Jednak kompulsywni użytkownicy pornografii internetowej silniej preferują nowe obrazy o charakterze seksualnym. Przyzwyczajają się do nich szybciej, co napędza poszukiwanie coraz to bardziej nowych obrazów seksualnych.
W chorobie Parkinsona leczenie zastępcze dopaminy (lewodopa, agonista i prekursor dopaminowy) wiąże się z kompulsywnymi zachowaniami seksualnymi i innymi zaburzeniami kontroli impulsów. Niektórzy pacjenci zgłaszali nietypowe kompulsywne używanie pornografi i wykazywali większą aktywność mózgu w odpowiedzi na obrazy seksualne, skolerowane ze zwiększonym pożądaniem seksualnym.
Czy uzależnienie od porno jest problemem? Niektóre źródła łączą pornografię ze wzrostem problemów z wydajnością seksualną i niskim pożądaniem seksualnym u mężczyzn poniżej 40 roku życia. Kiedy użytkownik porno powiązał swoje seksualne podniecenie z pornografią internetową, seks z pożądanymi prawdziwymi partnerami moze zostać zarejestrowany jako “niespełniający oczekiwań”, powodując odpowiedni spadek dopaminy i problemy z erekcją.
Inni nie zgadzają się, że nadmierna pornografia uzależnia, twierdząc, że na przykład oglądanie sportu (jak mecze koszykówki) prowadzi do podobnych procesów mózgowych u wielu osób.
Badania opisujące sensytyzację u osób uzależnionych od seksu lub użytkowników porno: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20.
Badania opisujące gorsze funkcjonowanie wykonawcze (hipofrontalność) lub zmienioną aktywność przedczołową u użytkowników porno / osób uzależnionych od seksu: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13.
Środki psychoaktywne
Istnieją substancje (naturalne i chemiczne) które są tzw. “drogą na skróty” w podnoszeniu poziomu dopaminy w mózgu. Wykazują one różne mechanizmy, dzięki którym również odczuwamy przyjemność: uwalniają one dopaminę z zakończeń nerwowych, pobudzają receptory dopaminowe, hamują rozkład dopaminy lub hamują jej wychwyt zwrotny – w dwóch ostatnich przypadkach mamy do czynienia z tzw. “recyclingiem” Taki mechanizm wyzwala np. amfetamina. Jeszcze inne substancje działają na zasadzie “hamulca hamulca“, tzn. hamują mechanizmy hamujące aktywację neuronów układu nagrody (np. alkohol oraz morfina).
Wzmacniające działanie substancji psychoaktywnych wynika nie tylko ze zwiększonej dawki dopaminy, ale również ze względu na szybki wzrost jej poziomu. A im szybszy wzrost tym bardziej intensywne efekty wzmacniające. Może to prowadzić do uszkodzenia lub zmniejszenia liczby receptorów dopaminy w pewnym okresie czasu, powodując zwiększone zapotrzebowanie na daną substancję.
Długotrwałe zażywanie substancji psychoaktywnych wydaje się być związane ze zmniejszeniem czynności dopaminy. Obniżenie receptorów dopaminergicznych D2 i uwalnianie dopaminy w prążkowiu u osób uzależnionych potwierdza tę hipotezę.
Wiele badań wykazało niską aktywność receptora D2/3 (w prążkowiu) u ludzi uzależnionych od addyktogenów.
W porównaniu z grupą kontrolną osoby zażywające kokainę wykazywały niższą dostepność receptorów D2/D3 w jądrze ogoniastym, skorupie i prążkowiu brzusznym.
U małp i gryzoni niska aktywność D2/3 skutkowała zwiększonym samodzielnym podawaniem kokainy. Natomiast w innym badaniu dominujące małpy z większą liczbą receptorów D2 same pobierały znacznie mniej kokainy w porównaniu z podrzędnymi zwierzętami.
Wydaje się zatem, że wysoki poziom receptorów D2 “chronił” dominujące małpy przed skutkami wzmacniającymi kokainę, co jest zgodne z danymi u zwierząt i ludzi.
Jak wyjaśniła psycholog Susan Weinschenk w artykule z 2009 r. – “dopamina neuroprzekaźnikowa nie powoduje u ludzi przyjemności, ale raczej jej poszukiwanie. Powoduje że pragniemy, pragniemy, szukamy i szukamy. Odpowiedzialnym za odczuwanie przyjemności jest układ opioidowy, jednak to układ dopaminowy jest silniejszy, dlatego szukamy więcej niż faktycznie jesteśmy z tego zadowoleni.
Zasadniczo, nasze zachowanie oraz wybory stają się coraz bardziej stronnicze w kierunku krótkoterminowych celów maksymalizacji przyjemności, tak jak w uzależnionym mózgu (jądro migdałowate, tylna brzuszna kora przedczołowa, prążkowie i jądro półleżące) zdala od długotrwałego dobrobytu i ogólnych celów maksymalizacji dobrostanu (normalnie zapewniane przez przednią korę przedczołową, hipokamp, kość grzbietowo-boczną przedczołową i korę obręczy przedniej).
Opracował: Kamil Bernatowicz
Literatura cytowana
1. Nestler E J, Hope B T, Widnell K L(1993) Neuron 11:995–1006, pmid:8274284
2. Berke J D, Hyman S E(2000) Neuron 25:515–532, pmid:10774721.
3. Nestler E J(2001) Nat Rev Neurosci 2:119–128, pmid:11252991.
4. Carlezon W A Jr, Thome J, Olson V G, Lane-Ladd S B, Brodkin E S, Hiroi N, Duman R S, Neve R L, Nestler E J(1998) Science 282:2272–2275, pmid:9856954
5. O’Donovan K J, Tourtellotte W G, Millbrandt J, Baraban J M(1999) Trends Neurosci 22:167–173
6. Mackler S A, Korutla L, Cha X Y, Koebbe M J, Fournier K M, Bowers M S, Kalivas P W(2000) J Neurosci 20:6210–6217
7. Morgan J I, Curran T(1995) Trends Neurosci 18:66–67, pmid:7537412
8. Young S T, Porrino L J, Iadarola M J(1991) Proc Natl Acad Sci USA 88:1291–1295,
9. Graybiel A M, Moratalla R, Robertson H A(1990) Proc Natl Acad Sci USA 87:6912–6916,
10. Hope B, Kosofsky B, Hyman S E, Nestler E J(1992) Proc Natl Acad Sci USA 89:5764–5768,
11. Kelz M B, Nestler E J(2000) Curr Opin Neurol 13:715–720, pmid:11148675
12. Koob G F, Sanna P P, Bloom F E(1998) Neuron 21:467–476, pmid:9768834.
13. Wise R A(1998) Drug Alcohol Dependence 51:13–22, pmid:9716927
14. Hope B T, Nye H E, Kelz M B, Self D W, Iadarola M J, Nakabeppu Y, Duman R S, Nestler E J(1994) Neuron 13:1235–1244
15. Nye H, Hope B T, Kelz M, Iadarola M, Nestler E J(1995) J Pharmacol Exp Ther 275:1671–1680,
16. Nye H E, Nestler E J(1996) Mol Pharmacol 49:636–645, pmid:8609891.
17. Moratalla R, Elibol B, Vallejo M, Graybiel A M(1996) Neuron 17:147–156
18. Pich E M, Pagliusi S R, Tessari M, Talabot-Ayer D, Hooft van Huijsduijnen R, Chiamulera C(1997) Science 275:83–86
19. Chen J S, Nye H E, Kelz M B, Hiroi N, Nakabeppu Y, Hope B T, Nestler E J(1995) Mol Pharmacol 48:880–889
20. Hiroi N, Brown J, Ye H, Saudou F, Vaidya V A, Duman R S, Greenberg M E, Nestler E J(1998) J Neurosci 18:6952–6962
21. Chen J, Kelz M B, Hope B T, Nakabeppu Y, Nestler E J(1997) J Neurosci 17:4933–4941,
22. Hiroi N, Brown J, Haile C, Ye H, Greenberg M E, Nestler E J(1997) Proc Natl Acad Sci USA 94:10397–10402
23. Fienberg A A, Hiroi N, Mermelstein P, Song W-J, Snyder G L, Nishi A, Cheramy A, O’Callaghan J P, Miller D, Cole D G, et al.(1998) Science 281:838–842
24. Hiroi N, Feinberg A, Haile C, Greengard P, Nestler E J(1999) Eur J Neurosci 11:1114–1118,
25. Greengard P, Allen P B, Nairn A C(1999) Neuron 23:435–447, pmid:10433257.
26. Bibb J A, Snyder G L, Nishi A, Yan Z, Meijer L, Fienberg A A, Tsai L H, Kwon Y T, Girault J A, Czernik A J, et al.(1999) Nature (London) 402:669–671
27. Chen J S, Kelz M B, Zeng G Q, Sakai N, Steffen C, Shockett P E, Picciotto M, Duman R S, Nestler E J(1998) Mol Pharmacol 54:495–503
28. Kelz M B, Chen J S, Carlezon W A, Whisler K, Gilden L, Beckmann A M, Steffen C, Zhang Y-J, Marotti L, Self S W, et al.(1999) Nature (London) 401:272–276
29. Dobrazanski P, Noguchi T, Kovary K, Rizzo C A, Lazo P S, Bravo R(1991) Mol Cell Biol 11:5470–5478,
30. Nakabeppu Y, Nathans D(1991) Cell 64:751–759, pmid:1900040
31. Yen J, Wisdom R M, Tratner I, Verma I M(1991) Proc Natl Acad Sci USA 88:5077–5081
32. White F J, Hu X-T, Zhang X-F, Wolf M E(1995) J Pharmacol Exp Ther 273:445–454,
33. Hyman S E(1996) Neuron 16:901–904, pmid:8630246
34. Kreek M J(1997) Pharmacol Biochem Behav 57:551–569
35. Shippenberg T S, Rea W(1997) Pharmacol Biochem Behav 57:449–455
36. Chen J S, Zhang Y J, Kelz M B, Steffen C, Ang E S, Zeng L, Nestler E J(2000) J Neurosci 20:8965–8971
37. Bibb J A, Chen J S, Taylor J R, Svenningsson P, Nishi A, Snyder G L, Yan Z, Sagawa Z K, Nairn A C, Nestler E J, et al.(2001) Nature (London) 410:376–380
38. Luscher C, Nicoll R A, Malenka R C, Muller D(2000) Nat Neurosci 3:545–550
39. Malinow R, Mainen Z F, Hayashi Y(2000) Curr Opin Neurobiol 10:352–357
40. Scannevin R H, Huganir R L(2000) Nat Rev Neurosci 1:133–141
41. Robinson, T. E. & Kolb, B. (1999) (1997) Eur. J. Neurosci.11, 1598–1604
42. Carey M, Smale S T(2000) Transcriptional Regulation in Eukaryotes (Cold Spring Harbor Lab. Press, Plainview, NY)
43. Spencer V A, Davie J R(1999) Gene 240:1–12, pmid:10564807
44. Shane W Kraus, Valerie Voon “Neurobiology of Compulsive Sexual Behavior: Emerging Science” Neuropsychopharmacology. 2016 Jan; 41(1): 385–386.
45. Brian Y. Park, Gary Wilson “Is Internet Pornography Causing Sexual Dysfunctions? A Review with Clinical Reports” Behav Sci (Basel). 2016 Sep; 6(3): 17
46. Rory C. Reid, Bruce N. Carpenter “Neuroscience research fails to support claims that excessive pornography consumption causes brain damage” Surg Neurol Int. 2011; 2: 64.
47. Christensen JF “Pleasure junkies all around! Why it matters and why ‘the arts’ might be the answer: a biopsychological perspective.” Proc Biol Sci. 2017 May 17;284(1854)
48. Volkow ND, Fowler JS “Dopamine in drug abuse and addiction: results of imaging studies and treatment implications.” Arch Neurol. 2007 Nov;64(11):1575-9.
49. Valerie Voon, Thomas B. Mole “Neural Correlates of Sexual Cue Reactivity in Individuals with and without Compulsive Sexual Behaviours” PLoS One. 2014; 9(7): e102419.
50. Mechelmans DJ1, Irvine M “Enhanced Attentional Bias towards Sexually Explicit Cues in Individuals with and without Compulsive Sexual Behaviours (2014)” PLoS One. 2014 Aug 25;9(8):e105476
51. Paula Banca et al. Novelty, conditioning and attentional bias to sexual rewards, Journal of Psychiatric Research (2016). DOI: 10.1016/j.jpsychires.2015.10.017
52. Analysis of “Sexual Desire, Not Hypersexuality, Is Related To Neurophysiological Responses Elicited by Sexual Images” (Steele et al., 2013)
53. Christian Laier&Matthias Brand “Empirical Evidence and Theoretical Considerations on Factors Contributing to Cybersex Addiction From a Cognitive-Behavioral View” Sexual Addiction & Compulsivity The Journal of Treatment & Prevention
Volume 21, 2014 – Issue 4
54. Brand M1, Laier C, Pawlikowski M “Watching pornographic pictures on the Internet: role of sexual arousal ratings and psychological-psychiatric symptoms for using Internet sex sites excessively.” Cyberpsychol Behav Soc Netw. 2011 Jun;14(6):371-7
55. Laier C, Schulte FP, Brand M. “Pornographic picture processing interferes with working memory performance.” J Sex Res. 2013;50(7):642-52.
56. Laier C, Pawlikowski M, Pekal J “Cybersex addiction: Experienced sexual arousal when watching pornography and not real-life sexual contacts makes the difference.” J Behav Addict. 2013 Jun;2(2):100-7
57. Laier C1, Pekal J, Brand M “Cybersex addiction in heterosexual female users of internet pornography can be explained by gratification hypothesis.”Cyberpsychol Behav Soc Netw. 2014 Aug;17(8):505-11
58. Snagowski J, Wegmann E, Pekal J “Implicit associations in cybersex addiction: Adaption of an Implicit Association Test with pornographic pictures.” Addict Behav. 2015 Oct;49:7-12. doi: 10.1016
59. Jan Snagowski and Matthias Brand “Symptoms of cybersex addiction can be linked to both approaching and avoiding pornographic stimuli: results from an analog sample of regular cybersex users” Front Psychol. 2015; 6: 653.
60. JOHANNES SCHIEBENER, CHRISTIAN LAIER “Getting stuck with pornography? Overuse or neglect of cybersex cues in a multitasking situation is related to symptoms of cybersex addiction” J Behav Addict. 2015 Mar; 4(1): 14–21.
61. Laier C, Pekal J, Brand M “Sexual Excitability and Dysfunctional Coping Determine Cybersex Addiction in Homosexual Males.” Cyberpsychol Behav Soc Netw. 2015 Oct;18(10):575-80.
62. Sina Wehrum-Osinsky, Dipl-Psych, Jan Schweckendiek “Altered Appetitive Conditioning and Neural Connectivity in Subjects With Compulsive Sexual Behavior” April 2016Volume 13, Issue 4, Pages 627–636
63. Jan Snagowski, Christian Laier, Theodora Duka, Matthias Brand “Subjective Craving for Pornography and Associative Learning Predict Tendencies Towards Cybersex Addiction in a Sample of Regular Cybersex Users” Sexual Addiction & Compulsivity The Journal of Treatment & Prevention Volume 23, 2016 – Issue 4
64. Paula Banca, Neil A. Harrison, Valerie Voon “Compulsivity Across the Pathological Misuse of Drug and Non-Drug Rewards” Front. Behav. Neurosci., 03 August 2016
65. Albery IP, Lowry J, Frings D “Exploring the Relationship between Sexual Compulsivity and Attentional Bias to Sex-Related Words in a Cohort of Sexually Active Individuals (2016)” Eur Addict Res. 2016 Oct 1;23(1):1-6.
66. Gola M, Wordecha M, Sescousse G “Can Pornography be Addictive? An fMRI Study of Men Seeking Treatment for Problematic Pornography Use.” Neuropsychopharmacology. 2017 Sep;42(10):2021-2031.
67. Christian Laier Matthias Brand “Mood changes after watching pornography on the Internet are linked to tendencies towards Internet-pornography-viewing disorder” Addictive Behaviors Reports Volume 5, June 2017, Pages 9-13
68. Rudolf Stark, Onno Kruse, Jan Snagowski “Predictors for (Problematic) Use of Internet Sexually Explicit Material: Role of Trait Sexual Motivation and Implicit Approach Tendencies Towards Sexually Explicit Material” Sexual Addiction & Compulsivity The Journal of Treatment & Prevention Volume 24, 2017 – Issue 3
69. Ferris Jabr “How the Brain Gets Addicted to Gambling” BEHAVIOR & SOCIETY, Scientific American
70. https://yourbrainonporn.com
71. https://lifehacker.com
72. http://alcoholrehab.com
73. https://www.sparringmind.com
74. https://www.soberrecovery.com
75. https://www.thecabinchiangmai.com
76. http://dopaminechallenge.com